Relation entre la teneur membranaire en n-3 HUFA et l’efficacité mitochondriale
 © Ifremer Stéphane Lesbats
© Ifremer Stéphane LesbatsLa relation entre la teneur membranaire en acides gras et l’efficacité mitochondriale diffère au sein et entre les teneurs alimentaires en oméga 3 à longue chaine.
Résumé
Une conséquence importante du changement climatique est la réduction de la production de nutriments essentiels à la base de la chaîne alimentaire marine : les acides gras polyinsaturés à longue chaîne de la série oméga-3 (n-3 HUFA). Cela peut avoir des conséquences dramatiques sur les consommateurs, comme les poissons, car ils ont une capacité limitée à synthétiser de novo les n-3 HUFA. Ces acides gras tels que l’acide docosahexaénoique (DHA, 22:6n-3) et l’acide eicosapentaénoique (EPA, 20:5n-3) sont essentiels pour la structure et la fonction des membranes biologiques. Il est de plus en plus évident que les poissons seront gravement affectés par la réduction de la disponibilité alimentaire des n-3 HUFA, mais les mécanismes sous-jacents restent méconnus. Les hypothèses sur les changements du fonctionnement des mitochondries en réponse à la disponibilité alimentaire en n-3 HUFA ont généralement négligé la production d’ATP, malgré son importance pour la capacité énergétique totale d’une cellule et, par conséquent, pour les performances de l’animal entier. Nous avons ici (i) quantifié la variation entre individus de l’efficacité mitochondriale (rapport ATP/O) du muscle et (ii) examiné sa relation avec la teneur en EPA et DHA de la membrane musculaire d’un poisson consommateur primaire, le mulet doré Chelon auratus, recevant un régime alimentaire riche ou pauvre en n-3 HUFA. Les mitochondries de poissons nourris avec un régime pauvre en n-3 HUFA présentaient un rapport ATP/O plus élevé que celles de poissons nourris avec un régime riche en n-3 HUFA. Cependant, l’efficacité mitochondriale variait environ deux fois entre les individus provenant du même traitement alimentaire, certains poissons consommant la moitié en oxygène et nutriment pour produire la même quantité d’ATP que leurs congénères issus d’un même régime alimentaire. Cette variation de l’efficacité mitochondriale entre les individus recevant la même alimentation était liée aux différences individuelles dans la composition en acides gras des membranes : un rapport ATP/O élevé était associé à une teneur élevée en EPA et DHA dans les membranes biologiques. Nos résultats mettent en évidence l’existence de différences interindividuelles dans l’efficacité mitochondriale et son importance potentielle pour expliquer la variation intra-spécifique en réponse aux changements dans la chaîne alimentaire.
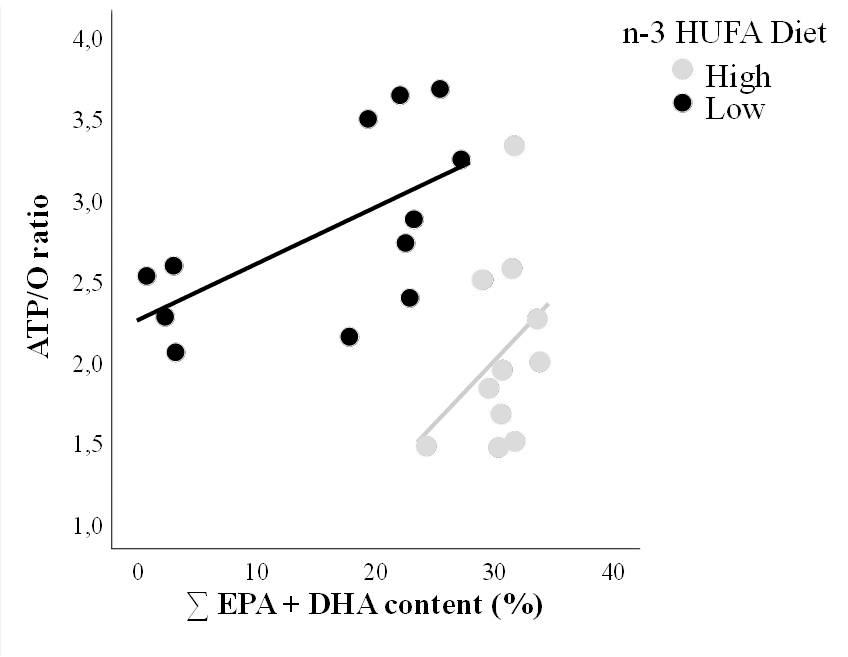
Figure 2 : L’efficacité à produire de l’ATP (ATP/O ratio) des mitochondries de mulet doré nourri avec un régime riche (High) ou pauvre (Low) en n-3 HUFA est correlée à la teneur en n-3 [acide docosahexaénoïque (DHA, 22:6n-3) et acide eicosapentaénoïque (EPA, 20:5n-3)] de ses phospholipides membranaires. Les individus dont les mitochondries sont plus efficaces à produire de l’ATP ont une teneur plus élevée en n-3 membranaire, et ce quelque soit leur régime alimentaire.
Réference: Salin K, Mathieu-Resuge M, Graziano N, Dubillot E, Le Grand F, Soudant P, Vagner M. 2020. The relationship between membrane fatty acid content and mitochondrial efficiency differs within- and between- omega-3 dietary treatments. Marine Environmental Research:105205.
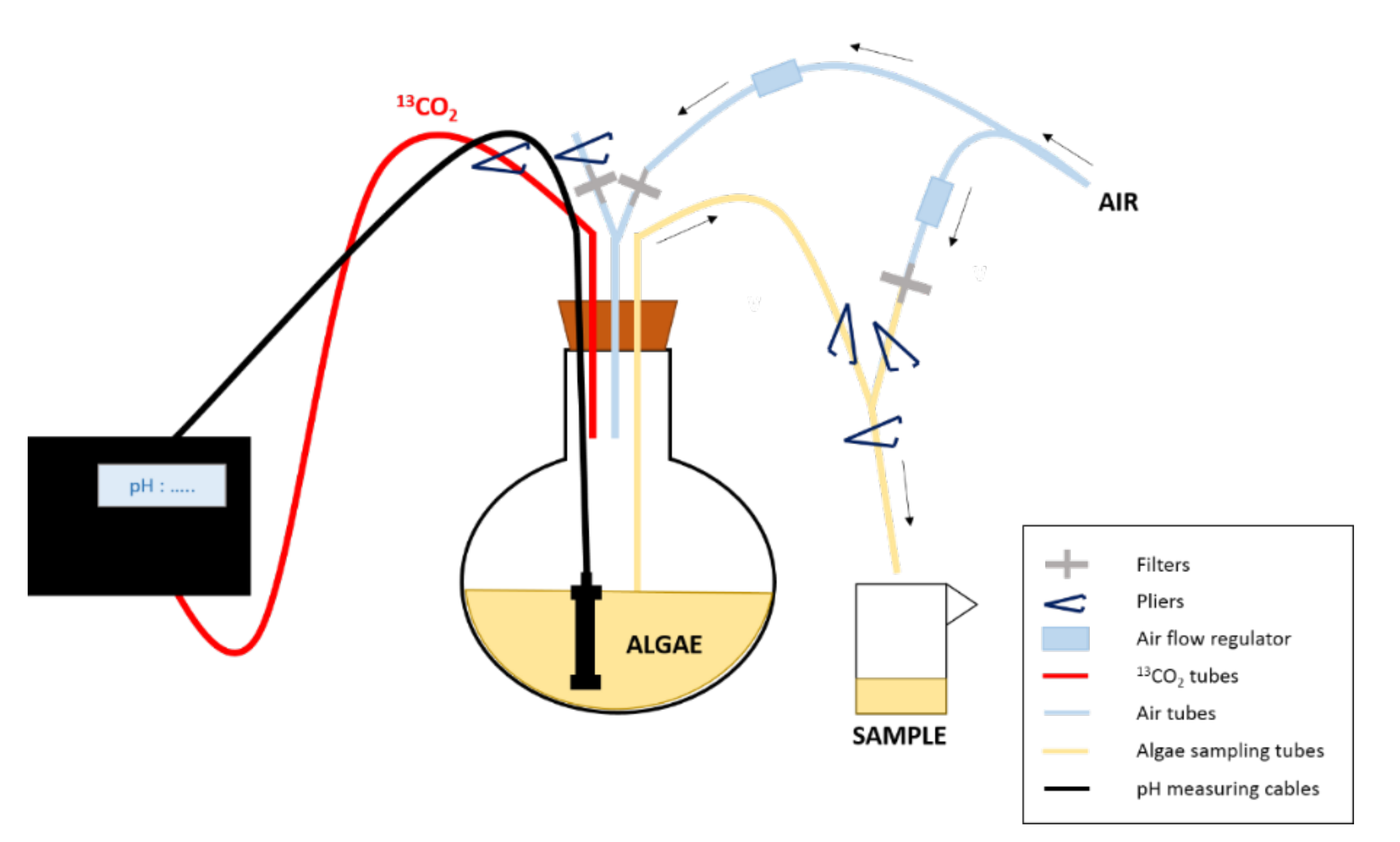
 Jeff Muir
Jeff Muir