Relationship between membrane n-3 HUFA content and mitochondrial efficiency

The relationship between membrane fatty acid content and mitochondrial efficiency differs within- and between- omega-3 dietary treatments
Abstract
An important, but underappreciated, consequence of climate change is the reduction in crucial nutrient production at the base of the marine food chain: the long-chain omega-3 highly unsaturated fatty acids (n-3 HUFA). This can have dramatic consequences on consumers, such as fish as they have limited capacity to synthesise n-3 HUFA de novo. The n-3 HUFA, such as docosahexaenoic acid (DHA, 22:6n-3) and eicosapentaenoic acid (EPA, 20:5n-3), are critical for the structure and function of all biological membranes. There is increasing evidence that fish will be badly affected by reductions in n-3 HUFA dietary availability, however the underlying mechanisms remain obscure. Hypotheses for how mitochondrial function should change with dietary n-3 HUFA availability have generally ignored ATP production, despite its importance to a cell’s total energetics capacity, and in turn, whole-animal performance. Here we (i) quantified individual variation in mitochondrial efficiency (ATP/O ratio) of muscle and (ii) examined its relationship with content in EPA and DHA in muscle membrane of a primary consumer fish, the golden grey mullet Chelon auratus, receiving either a high or low n-3 HUFA diet. Mitochondria of fish fed on the low n-3 HUFA diet had higher ATP/O ratio than those of fish maintained on the high n-3 HUFA diet. Yet, mitochondrial efficiency varied up about 2-fold among individuals on the same dietary treatment, resulting in some fish consuming half the oxygen and energy substrate to produce the similar amount of ATP than conspecific on similar diet. This variation in mitochondrial efficiency among individuals from the same diet treatment was related to individual differences in fatty acid composition of the membranes: a high ATP/O ratio was associated with a high content in EPA and DHA in biological membranes. Our results highlight the existence of interindividual differences in mitochondrial efficiency and its potential importance in explaining intraspecific variation in response to food chain changes.
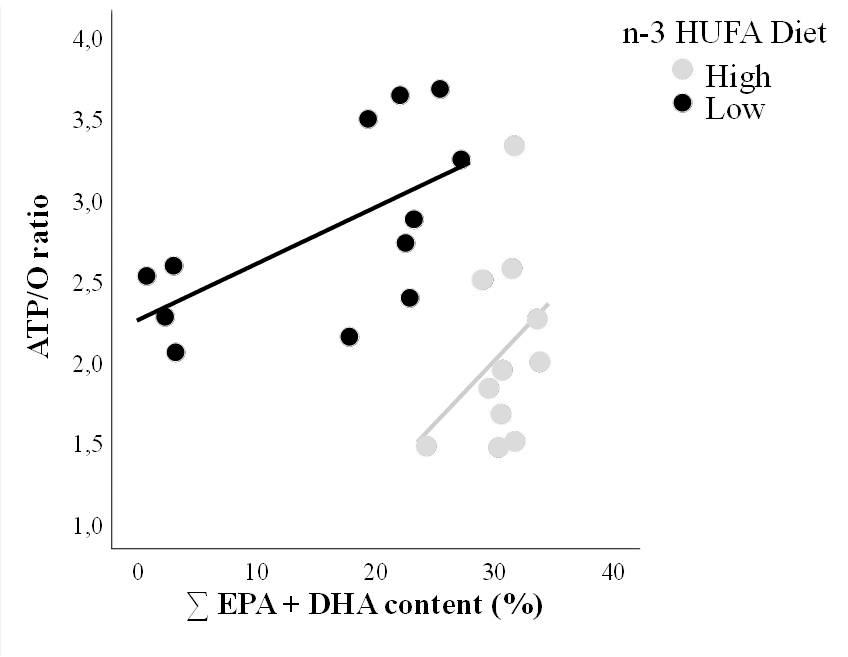
Figure 2: A golden grey mullet’s mitochondrial efficiency of ATP production under Low and High n-3 HUFA diet is related to the n-3 content (docosahexaenoic acid (DHA, 22:6n-3) and eicosapentaenoic acid (EPA, 20:5n-3)) of its membrane phospholipids, with individuals that have mitochondria with higher ability to make ATP had higher membrane n-3 content.
Reference
Salin K, Mathieu-Resuge M, Graziano N, Dubillot E, Le Grand F, Soudant P, Vagner M. 2020. The relationship between membrane fatty acid content and mitochondrial efficiency differs within- and between- omega-3 dietary treatments. Marine Environmental Research:105205.