Identification des voies de synthèse des acides gras polyinsaturés dans le dinophyte toxique Alexandrium minutum à l’aide du marquage au 13C
Résumé :
Les voies de synthèse responsables de la production des acides gras polyinsaturés 22:6n-3 et 20:5n-3 ont été étudiées dans le Dinophyte Alexandrium minutum. Le but de ce travail était de suivre l’incorporation progressive d’un marqueur isotopique (13CO2) dans 11 acides gras pour mieux comprendre les voies de synthèse des acides gras dans A. minutum. La croissance des dinophytes a été suivie pendant 54 h en utilisant un échantillonnage à haute fréquence. A. minutum a présenté une croissance en deux phases. Une phase de latence a été observée pendant les 30 premières heures de développement et a été associée à l’enkystement temporaire probable des cellules Dinophyte. Une phase de croissance exponentielle a ensuite été observée après t30. A. minutum a rapidement incorporé du 13C dans 22:6n-3, qui a fini par être l’acide gras polyinsaturé (AGPI) le plus enrichi en 13C dans cette expérience, avec un enrichissement atomique en 13C supérieur à celui de 18:4n-3, 18:5n-3, 20:5n-3 et 22:5n-3. Dans l’ensemble, l’enrichissement atomique en 13C était inversement proportionnel au nombre de carbones dans l’AGPI n-3. Les AGPI C18, 18:4n-3, et 18:5n-3, étaient en effet parmi les AG les moins enrichis en 13C au cours de cette expérience. On a supposé qu’ils étaient produits par la voie des AGPI n-3. Cependant, ils n’ont pas pu être allongés ou désaturés davantage pour produire des AGPI n-3 en C20-C22, car les AE des AGPI n-3 en C18 étaient inférieurs à ceux des AGPI n-3 en C20-C22. Ainsi, l’enrichissement atomique particulièrement élevé de 22:6n-3 (55,8% et 54,9% en lipides neutres (NL) et en lipides polaires (PL), respectivement) nous a conduit à émettre l’hypothèse que cet AGPI majeur était synthétisé par une voie de synthèse de la polykétide-synthase (PKS) indépendante de l’O2. Une autre PKS parallèle, indépendante de celle qui conduit à 22:6n-3, était également censée produire 20:5n-3. L’ordre inverse de l’enrichissement atomique en 13C pour les AGPI n-3 a également été suspecté d’être lié à l’éventuelle β-oxydation des AGPI n-3 à longue chaîne se produisant lors de l’encyssement d’A. minutum.
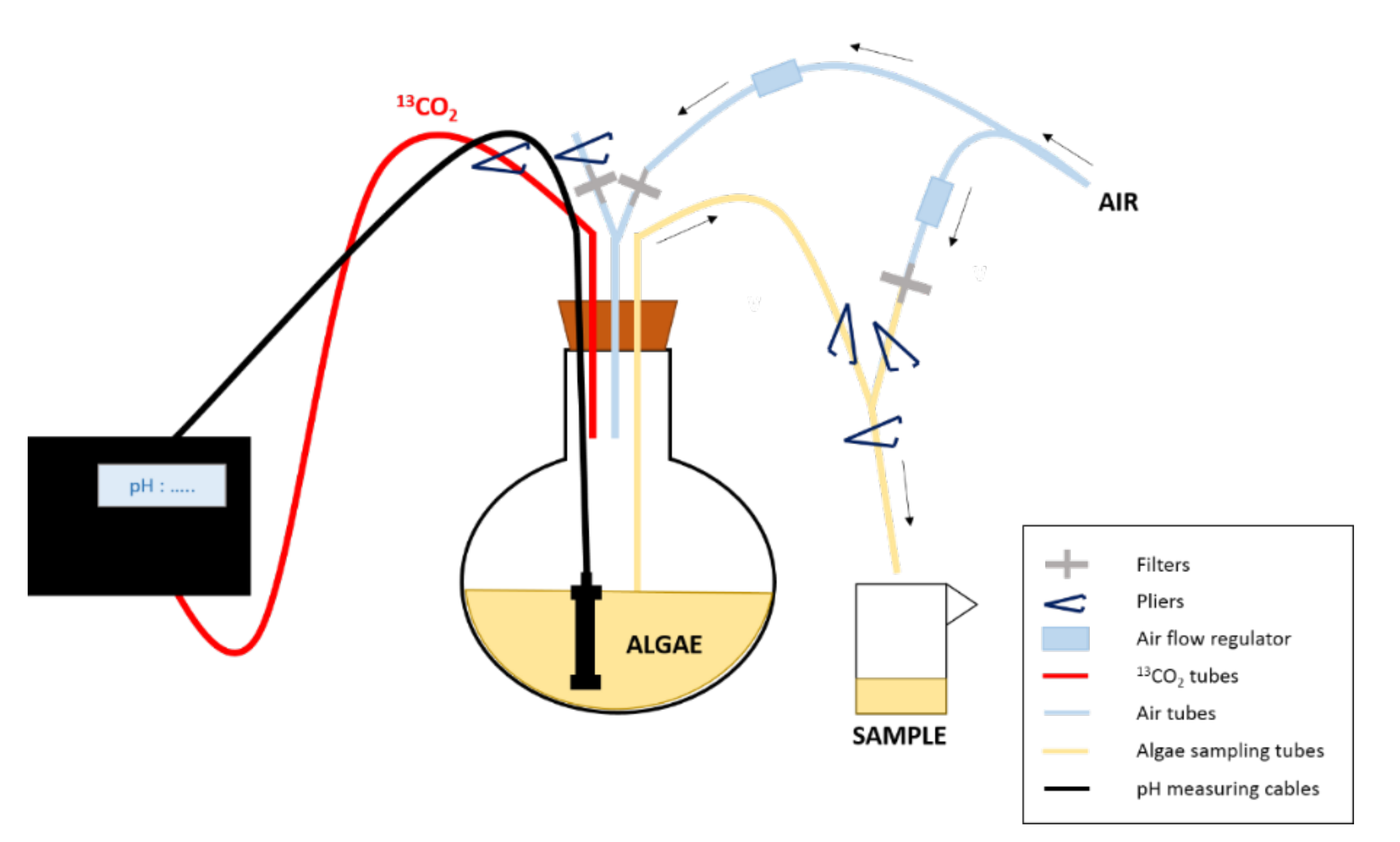
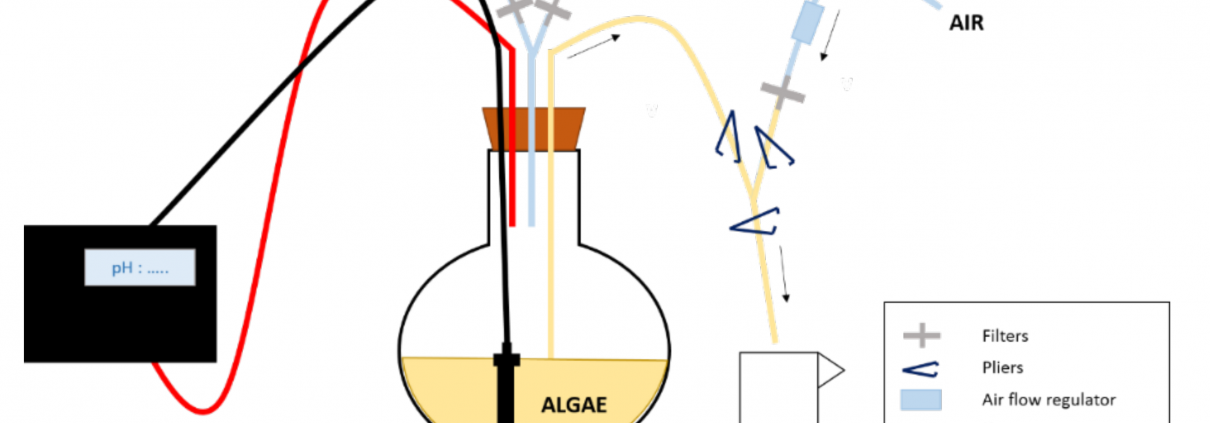
Figure 1 : Conception expérimentale de l’expérience d’enrichissement. Le 13CO2 est fourni à la culture en fonction de son pH. Pour prélever les algues, on utilise une pince pour fermer/ouvrir les tubes/voies nécessaires pour mettre d’abord le ballon sous pression, puis permettre le prélèvement et enfin rincer les tubes après le prélèvement.
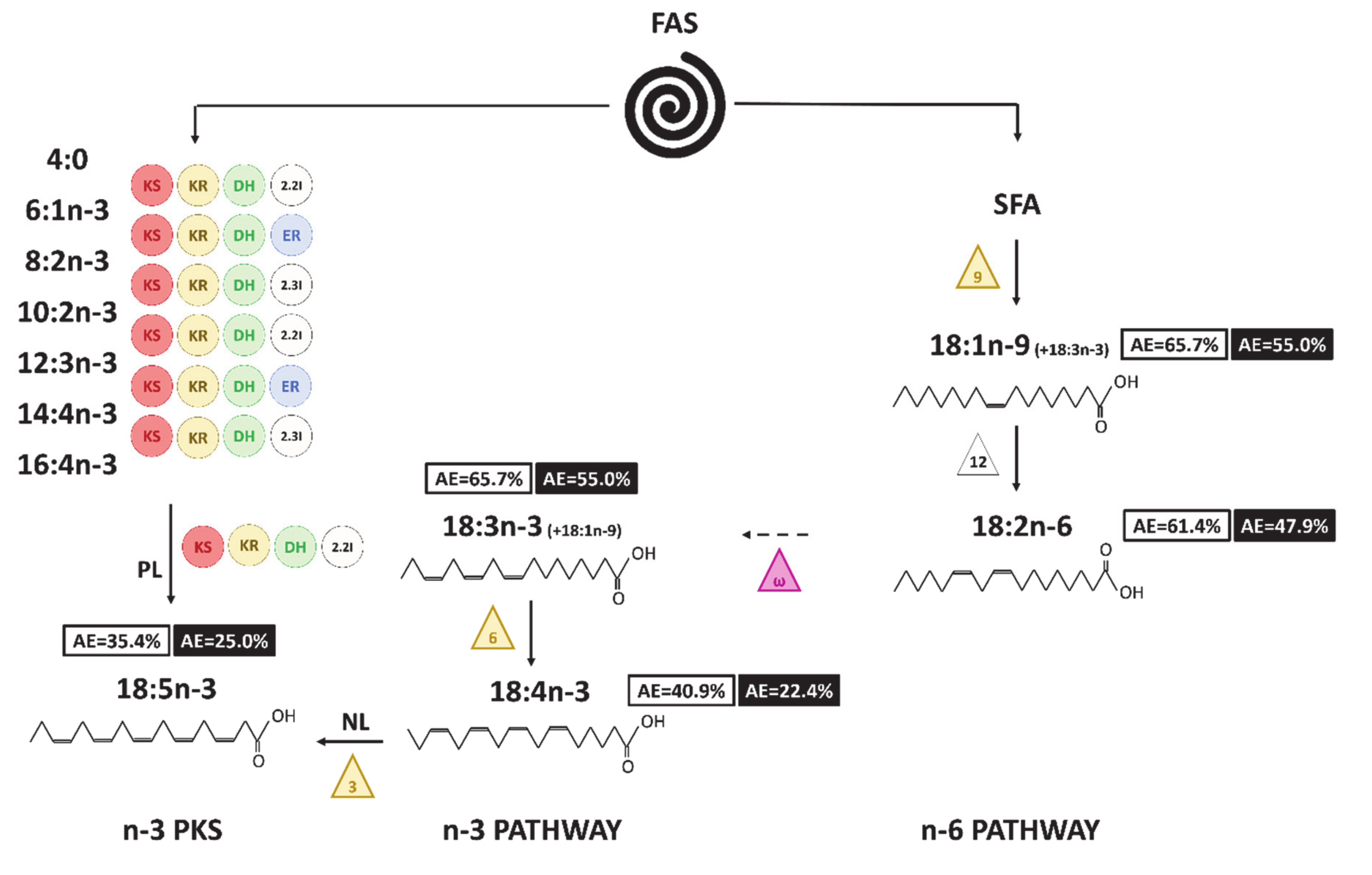
Figure 8 : Hypothèse de voies de production d’Acide Gras (AG) C18 chez A. minutum. Les chiffres dans les cases correspondent à la valeur finale de l’AG (en blanc pour les lipides neutres et en noir pour les lipides polaires). Les triangles symbolisent les désaturases (extrémité avant en jaune et extrémité méthyle en violet), les cercles les enzymes impliquées dans la voie PKS (KS : 3-ketoacyl synthase ; KR : 3-ketoacyl-ACP-reductase ; DH : déshydrase ; 2.2I : 2-trans, 2-cis isomérase ; 2.3I : 2-trans, 3-cis isomérase ; ER : enoyl réductase). Les voies avec les flèches en pointillés semblent peu probables ou ne peuvent être prouvées avec la dynamique de l’enrichissement. NL et PL écrits sur les voies permettant la synthèse de 18:5n-3 indiquent la fraction considérée pour chaque voie.
Conclusions :
La synthèse des acides gras chez le Dinophyte Alexandrium minutum a suivi différentes voies. La voie PKS semble être un processus de synthèse particulièrement rapide, responsable du fort enrichissement et de la production de l’acide gras polyinsaturé 22:6n-3. Le 22:6n-3 semble avoir un rôle central dans le maintien d’un bon état physiologique, y compris pendant l’enkystement. Dans notre étude, nous avons supposé que la phase de latence observée reflétait la possibilité d’un enkystement temporaire chez A. minutum. Elle était caractérisée par une diminution significative de la proportion de lipides neutres, correspondant à la consommation des lipides de réserve. Le 22:6n-3 pourrait être dégradé au cours de ce processus, et ses produits de dégradation pourraient être impliqués dans la re-synthèse du triacylglycérol lors de l’excystration des algues. La dynamique d’enrichissement des AGPI C18 a révélé qu’il est peu probable qu’ils soient impliqués dans les étapes de désaturation et d’allongement ultérieures des AGPI n-3 C20-C22. Ils semblent être les produits finaux de la voie n-3 classique. L’enrichissement atomique 18:5n-3 rend possible son origine à partir de la désaturation du 18:4n-3, de la dégradation d’acides gras plus longs tels que le 20:5n-3, ou de la voie PKS. On a supposé que ces AGPI en C18 jouent un certain rôle chez A. minutum, par exemple dans sa toxicité. Des études supplémentaires sont nécessaires pour mieux limiter la voie de synthèse des AGPI chez A. minutum, et surtout pour démontrer davantage l’implication de la voie PKS après la synthèse 22:6n-3 et 20:5n-3.
Réference :