Profondeur de recherche de nourriture des requins pélagiques : Les isotopes stables du mercure nous éclairent
 Mark Conlin/NMFS
Mark Conlin/NMFSRésumé
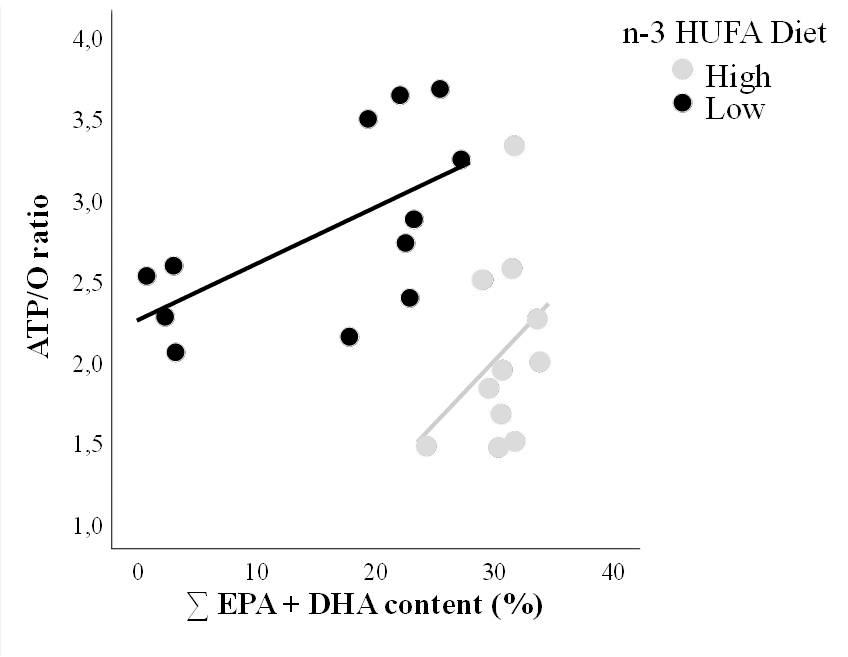
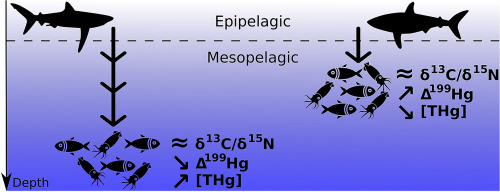
Le déclin des populations de requins dans l’océan mondial affecte la structure et la fonction des écosystèmes de manière imprévisible et de nouvelles informations écologiques sont aujourd’hui nécessaires pour mieux comprendre le rôle des requins dans leurs habitats. En particulier, la caractérisation des modèles de recherche de nourriture est cruciale pour comprendre et prévoir l’évolution de la dynamique entre les requins et leurs proies. De nombreuses espèces de requins utilisent la zone mésopélagique comme principale zone de recherche de nourriture, mais on ignore encore dans quelle mesure les différents requins pélagiques utilisent cet habitat. Afin d’illustrer la dimension verticale de leur écologie trophique, nous avons utilisé les isotopes stables du mercure dans le muscle de trois espèces de requins pélagiques (le requin bleu Prionace glauca, le requin-taupe bleu Isurus oxyrinchus et le requin-marteau lisse Sphyrna zygaena) de la région du Pacifique nord-est. Les valeurs Δ199Hg, allant de 1,40 à 2,13‰ chez les requins, suggéraient un régime alimentaire principalement basé sur des proies mésopélagiques dans des habitats océaniques. Nous avons en outre utilisé les isotopes stables du carbone et de l’azote (δ13C, δ15N) seuls ou en combinaison avec les valeurs Δ199Hg, pour évaluer le partage des ressources entre les trois espèces de requins. L’ajout de Δ199Hg a entraîné une diminution des estimations de chevauchement trophique par rapport à celles basées sur δ13C/δ15N seuls, ce qui démontre que la modélisation multi-isotope est nécessaire pour une description trophique précise des trois espèces. Principalement, cela révèle qu’ils s’alimentent à des profondeurs moyennes différentes et que le partage des ressources est principalement exprimé par la dimension verticale au sein des assemblages de requins pélagiques. Parallèlement, la concentration de mercure total (THg) dans les muscles diffère selon les espèces et augmente avec la profondeur d’alimentation. Dans l’ensemble, cette étude souligne le rôle clé de la zone mésopélagique pour les espèces de requins qui s’alimentent parmi d’importants gradients de profondeur et rapporte de nouvelles informations écologiques sur la compétition trophique en utilisant les isotopes du mercure. Elle suggère également que la profondeur de recherche de nourriture peut jouer un rôle central dans les différences entre les THg musculaires des espèces de requins cooccurrentes de niveau trophique élevé.
Résumé graphique
Points forts
– δ13C, δ15N, Δ199Hg et δ202Hg ont été déterminés chez trois espèces de requins pélagiques.
– Les isotopes de Hg suggèrent que ces espèces de requins se nourrissent de proies mésopélagiques.
– δ13C et δ15N surestiment les chevauchements entre les niches trophiques.
– Les différences de profondeur de recherche de nourriture expliquent mieux le partage des ressources.
– La profondeur de recherche de nourriture influence le niveau de contamination au mercure.
Référence
Lucien Besnard, Gaël Le Croizier, Felipe Galván-Magaña,David Point,Edouard Kraffe, James Ketchum, Raul Octavio Martinez Rincon, Gauthier Schaal. Foraging depth depicts resource partitioning and contamination level in a pelagic shark assemblage: Insights from mercury stable isotopes. Environmental Pollution, Volume 283, 2021, 117066. ISSN 0269-7491. https://doi.org/10.1016/j.envpol.2021.117066.

 © Ifremer Stéphane Lesbats
© Ifremer Stéphane Lesbats