Influence des ligands forts du fer sur la capacité d’oxydation de l’eau des nuages

Résumé
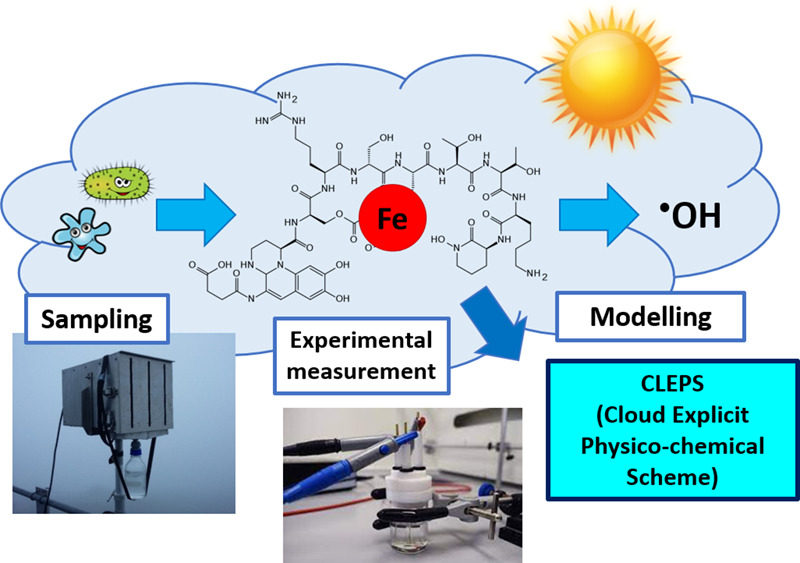
Le fer (Fe) joue un double rôle dans la chimie atmosphérique : il est impliqué dans la réactivité chimique et photochimique et sert de micronutriment pour les micro-organismes dont on a récemment montré qu’ils produisaient des ligands organiques puissants. Ces ligands contrôlent la réactivité, la mobilité, la solubilité et la spéciation du Fe, ce qui a un impact potentiel sur la biodisponibilité du Fe et la capacité oxydante de l’eau des nuages.
Dans ce travail, les concentrations de ligands liant le Fe et les constantes de stabilité conditionnelle ont été mesurées expérimentalement pour la première fois par la technique CLE-ACSV (Competitive Ligand Exchange-Adsorptive Cathodic Stripping Voltammetry) dans des échantillons d’eau trouble collectés au puy de Dôme (France). Les constantes de stabilité conditionnelles, qui indiquent la force des complexes Fe-ligand, sont plus élevées que celles considérées jusqu’à présent dans la chimie des nuages (principalement Fe-oxalate). Pour comprendre l’effet de la complexation du Fe sur la réactivité de l’eau des nuages, nous avons utilisé le modèle CLEPS de chimie des nuages. Selon les résultats du modèle, nous avons constaté que la complexation du Fe a un impact sur le taux de formation des radicaux hydroxyles : contrairement à nos attentes, la complexation du Fe par des ligands organiques naturels a conduit à une augmentation de la production de radicaux hydroxyles. Ces résultats ont un impact important sur la chimie des nuages et le cycle global du fer.
Points forts
- 95% du fer est complexé par des ligands organiques forts, probablement produits par des micro-organismes.
- Les constantes de stabilité des complexes du fer sont beaucoup plus élevées que celles utilisées dans la chimie des nuages.
- La présence de ligands organiques forts induit une augmentation de la production de radicaux hydroxyles.
- L’analyse des sources et des puits du point radicalaire HO2/O2 a mis en évidence que le fer complexé n’épuise pas le point radicalaire HO2/O2.
Référence
Aridane G. González, Angelica Bianco, Julia Boutorh, Marie Cheize, Gilles Mailhot, Anne-Marie Delort, Hélène Planquette, Nadine Chaumerliac, Laurent Deguillaume, Geraldine Sarthou. Influence of strong iron-binding ligands on cloud water oxidant capacity, Science of The Total Environment, Volume 829, 2022, 154642, ISSN 0048-9697, https://doi.org/10.1016/j.scitotenv.2022.154642.
 (C) M. Lutier
(C) M. Lutier


 Paul Einerhand
Paul Einerhand