Dernières avancées dans les interactions bivalve-microbiote pour la prévention des maladies en aquaculture

Résumé :
Chez les bivalves, aucun rôle fonctionnel précis du microbiote n’a encore été identifié, bien que de nombreuses publications suggèrent qu’il pourrait être impliqué dans la nutrition ou l’immunité de son hôte. Dans le contexte du changement climatique, des approches intégratives au carrefour des disciplines ont été développées pour explorer le système environnement-hôte-pathogène-microbiote. Nous tentons ici de synthétiser les travaux sur (1) les méthodologies actuelles d’analyse du microbiote des bivalves, (2) la comparaison du microbiote entre les espèces, entre les compartiments de l’hôte et son habitat environnant, (3) la manière dont le microbiote des bivalves est régi par les facteurs environnementaux et la génétique de l’hôte et (4) la manière dont les micro-organismes associés à l’hôte agissent comme un tampon contre les pathogènes et/ou favorisent la guérison, et pourraient ainsi jouer un rôle dans la prévention des maladies ou des mortalités.
Résumé graphique :
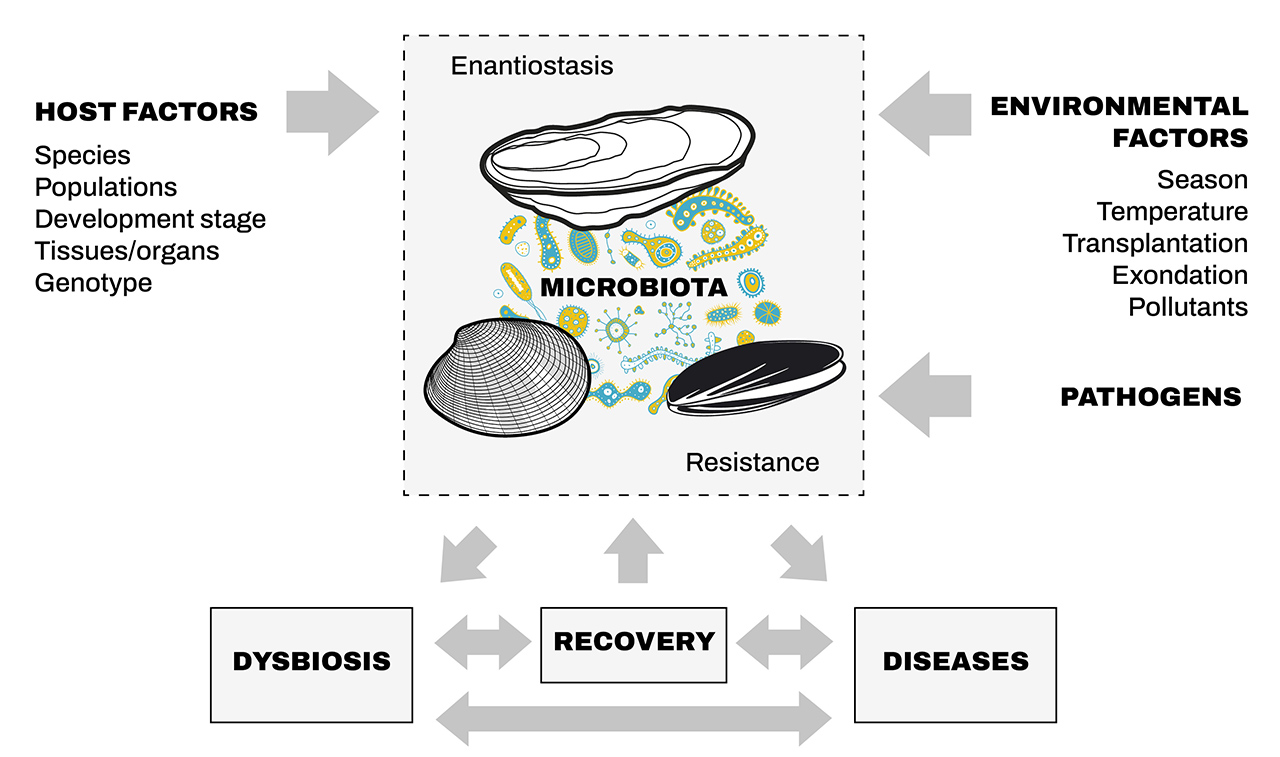
Figure 2. Facteurs de l’hôte et de l’environnement agissant sur le microbiote bivalve.
Il a été démontré que certains de ces facteurs, seuls ou combinés, entraînent une dysbiose, des maladies et une guérison.
Points forts :
- Les technologies de séquençage à haut débit ont permis de mener des recherches sur les interactions entre les bivalves et le microbiote.
- Les interactions bivalve-microbiote dépendent de facteurs de l’hôte imbriqués les uns dans les autres.
- L’interaction du microbiote avec le milieu environnant est importante pour la santé des bivalves.
- Des études récentes soulignent l’implication du microbiote des huîtres dans la résistance aux maladies.
- Les études sur le microbiome fourniront de nouvelles stratégies pour améliorer l’aquaculture des bivalves.
Référence :
Christine Paillard, Yannick Gueguen, K Mathias Wegner, David Bass, Alberto Pallavicini, Luigi Vezzulli, Isabelle Arzul. Recent advances in bivalve-microbiota interactions for disease prevention in aquaculture. Current Opinion in Biotechnology, Volume 73, 2022, Pages 225-232. ISSN 0958-1669. https://doi.org/10.1016/j.copbio.2021.07.026.
 Paul Einerhand
Paul Einerhand
 Mark Conlin/NMFS
Mark Conlin/NMFS

 © Ifremer Stéphane Lesbats
© Ifremer Stéphane Lesbats