Evidence that Pacific tuna mercury levels are driven by marine methylmercury production and anthropogenic inputs

Abstract
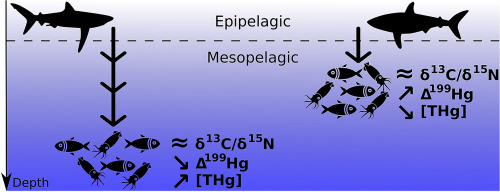
Pacific Ocean tuna is among the most-consumed seafood products but contains relatively high levels of the neurotoxin methylmercury. Limited observations suggest tuna mercury levels vary in space and time, yet the drivers are not well understood. Here, we map mercury concentrations in skipjack tuna across the Pacific Ocean and build generalized additive models to quantify the anthropogenic, ecological, and biogeochemical drivers. Skipjack mercury levels display a fivefold spatial gradient, with maximum concentrations in the northwest near Asia, intermediate values in the east, and the lowest levels in the west, southwest, and central Pacific. Large spatial differences can be explained by the depth of the seawater methylmercury peak near low-oxygen zones, leading to enhanced tuna mercury concentrations in regions where oxygen depletion is shallow. Despite this natural biogeochemical control, the mercury hotspot in tuna caught near Asia is explained by elevated atmospheric mercury concentrations and/or mercury river inputs to the coastal shelf. While we cannot ignore the legacy mercury contribution from other regions to the Pacific Ocean (e.g., North America and Europe), our results suggest that recent anthropogenic mercury release, which is currently largest in Asia, contributes directly to present-day human mercury exposure.
Graphical abstract
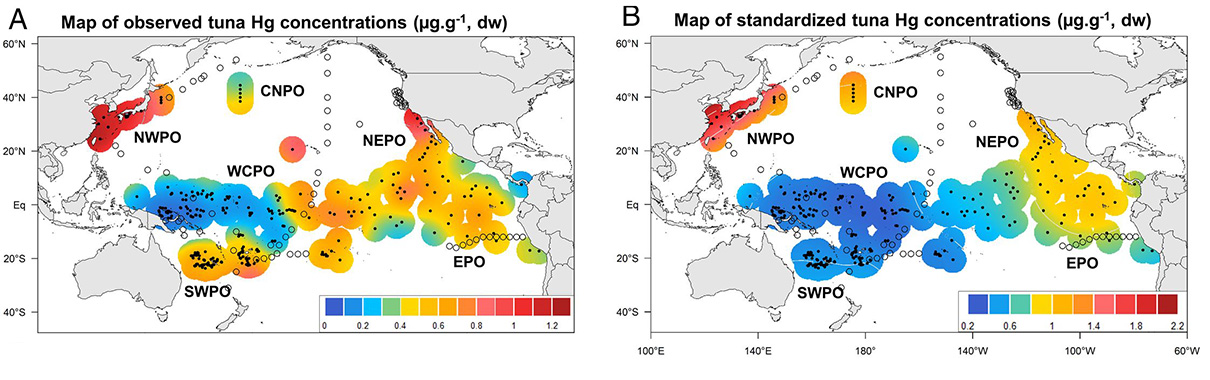
Spatial variability of skipjack mercury concentrations. Smoothed spatial contour maps of (A) observed and (B) standardized Hg concentrations (micrograms ⋅ grams−1, dw) in skipjack white muscle samples from the Pacific Ocean. The black dots represent the location of skipjack samples. Ocean areas correspond to the sample origin: NWPO, CNPO, NEPO, EPO, SWPO, and WCPO. The transparent dots represent the location of seawater samples with available and published MeHg data
Highlights
Humans are exposed to toxic methylmercury mainly by consuming marine fish. New environmental policies under the Minamata Convention rely on a yet-poorly-known understanding of how mercury emissions translate into fish methylmercury levels. Here, we provide the first detailed map of mercury concentrations from skipjack tuna across the Pacific. Our study shows that the natural functioning of the global ocean has an important influence on tuna mercury concentrations, specifically in relation to the depth at which methylmercury concentrations peak in the water column. However, mercury inputs originating from anthropogenic sources are also detectable, leading to enhanced tuna mercury levels in the northwestern Pacific Ocean that cannot be explained solely by oceanic processes.
Reference
Anaïs Médieu, David Point, Takaaki Itai, Hélène Angot, Pearse J. Buchanan, Valérie Allain, Leanne Fuller, Shane Griffiths, David P. Gillikin, Jeroen E. Sonke, Lars-Eric Heimbürger-Boavida, Marie-Maëlle Desgranges, Christophe E. Menkes, Daniel J. Madigan, Pablo Brosset, Olivier Gauthier, Alessandro Tagliabue, Laurent Bopp, Anouk Verheyden, Anne Lorrain. Evidence that Pacific tuna mercury levels are driven by marine methylmercury production and anthropogenic inputs. Proceedings of the National Academy of Sciences Jan 2022, 119 (2) e2113032119; DOI: 10.1073/pnas.2113032119